Este post é uma tradução do artigo que postei recentemente em meu blog anglófono, Montogue Quiz. Veja o post original aqui.
1. Introdução
Cianobactérias, os componentes mais numerosos do fitoplâncton, produzem toxinas e compostos malcheirosos que podem prejudicar a qualidade da água de ambientes lacustres e, de quebra, impedir seu uso em aplicações de importância econômica como irrigação, aquicultura e recreação. Sob condições ideais, cianobactérias podem multiplicar-se exponencialmente e produzir as chamadas florações de algas nocivas.
Há certa controvérsia acerca dos fatores que determinam a dominância cianobacteriana em ambientes de água doce. Sabendo que muitas algas florantes são capazes de fixar gás nitrogênio, alguns pesquisadores sugeriram que cianobactérias devem dominar sob razões N:P (nitrogênio/fósforo) reduzidas. Essa hipótese foi endossada em Smith (1983), que, com base em observações obtidas em 20 lagos do norte europeu, concluiu que o N:P ambiente de fato deve influenciar a dominância cianobacteriana. Não obstante, Trimbee e Prepas (1987) reanalisaram os dados de Smith e concluíram que concentrações de nutrientes (ou seja, P total ou N total) são melhores preditores de abundância cianobacteriana do que a razão N:P. Do mesmo modo, Downing et al. (2001) e Wagner e Adrian (2009) avaliaram diversas variáveis e concluíram que a razão N:P pode não ser um bom preditor de abundância cianobacteriana.
Ademais, pesquisas mais recentes indicam que não devemos atribuir a mesma dependência em gradientes nutricionais a todas as espécies de algas procariotas. Dolman et al. (2012) observaram que nove táxons de cianobactérias diferem substancialmente em suas respostas a variações em N total ou P total; é interessante observar que Dolman e seu grupo verificaram que dois táxons de Nostocales, que são fixadoras de nitrogênio, obtiveram biovolumes maiores em lagos com alta concentração de N e baixa concentração de P, ao contrário do que seria esperado. Há, ainda, um crescente corpo de pesquisa acerca da resposta de cianobactérias a outros fatores, como pH e temperatura (Kosten et al., 2012). Por exemplo, embora cianobactérias em lagos polimíticos possam não beneficiar-se diretamente com águas mais quentes, é certo que estas tiram proveito de períodos de estratificação mais extensos e maior oferta de nutrientes no hipolímnio, especialmente fósforo (Wagner e Adrian, 2009). Isso sugere que florações de algas em ambientes lacustres podem tornar-se mais comuns e intensas com o aquecimento climático.
De fato, há evidências de que florações de algas estão tornando-se mais frequentes em escala global; por exemplo, uma análise de pigmentos cianobacterianos em sedimentos de mais de 100 lagos norte-americanos e europeus demonstrou que populações de cianobactérias aumentaram substancialmente em 58% dos lagos desde a revolução industrial, e tal crescimento tem acelerado desde meados da década de 1940 (Taranu et al., 2015).
À medida que a eutroficação de ambientes lacustres se intensificou na segunda metade do século XX, stakeholders responderam com diversas técnicas de controle e mitigação. No post de hoje, discutimos brevemente cinco tais técnicas: tratamento químico, dragagem de sedimentos, aeração/oxigenação hipolimnética, biomanipulação e ultrassom. Algumas dessas técnicas (por exemplo, uso de sulfato de cobre) são usadas para combater florações diretamente, ao passo que outras (por exemplo, oxigenação artificial) são mais comumente utilizadas pós-eutroficação, com o intento de melhorar a qualidade da água.
2. Tratamento químico: sulfato de cobre, palha de cevada, peróxido de hidrogênio
2.1. Sulfato de cobre
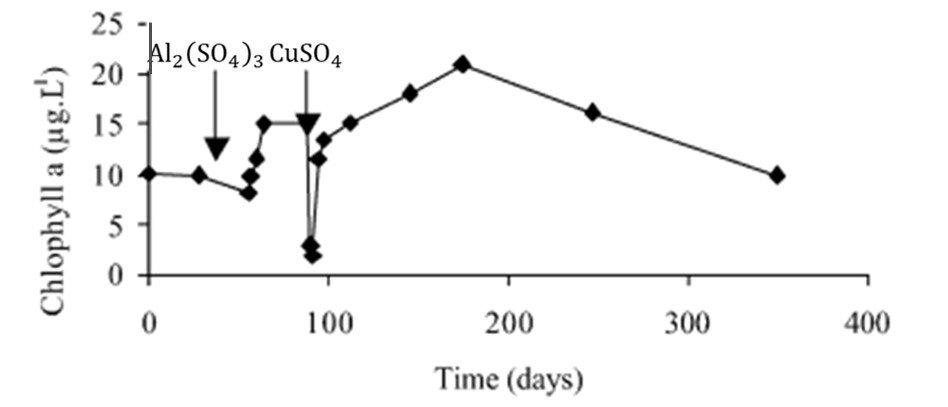
Usado na gestão de lagos há mais de 100 anos, sulfato de cobre (CuSO4) foi um dos primeiros algicidas. Uma de suas principais vantagens é a facilidade de aplicação, pois CuSO4 é vendido em diversas formulações (geralmente granular ou cristalino) e pode ser aplicado de várias formas. Concentrações práticas divulgadas na literatura variam de 25 a 1000 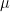
Figura 1. Clorofila-a versus tempo no Lago Courtille, França. As setas indicam as datas de aplicação de sulfato de alumínio e sulfato de cobre.
Ademais, sulfato de cobre não é bioespecífico e constitui uma ameaça para plantas aquáticas, insetos aquáticos e zooplâncton. A toxicidade de cobre para biota aquática em condições de laboratório decresce na ordem crustáceos (10
2.2. Herbicidas
No passado, herbicidas foram uma escolha óbvia de algicida porque supostamente poderiam controlar cianobactérias e algas eucariotas sem afetar formas de vida heterotróficas. Dois herbicidas comumente adotados no controle de algas são diuron e endothall. Diuron é um inibidor de fotossíntese; apesar de ser eficaz no controle de fitoplâncton, apresenta alta persistência em sedimentos (até 1 ano) e pode ser ambientalmente degradado a 3,4-dicloroanilina, uma substância genotóxica. Endothall, por sua vez, interfere na síntese de RNA; é altamente seletivo para cianobactérias, mas apresenta caráter tóxico para alguns invertebrados aquáticos. Desenvolvimento de resistência a endothall em populações algais já foi documentado (Prosecka et al., 2009).
2.3. Palha de cevada
Em vista do potencial tóxico de agentes sintéticos como diuron e endothall, alguns stakeholders têm advogado o uso de produtos naturais que apresentam efeito algicida. O produto do gênero mais amplamente pesquisado é palha de cevada, que tem sido considerada no controle de florações desde a década de 1990. Experimentos conduzidos no Reino Unido indicam que a aplicação de palha de cevada em concentrações de 25 – 50 g m–3 pode suprimir o crescimento de várias algas planctônicas por 3 – 6 meses, a partir de 12 dias a 2 meses após a aplicação (Everall e Lees, 1997). Islami e Filizadeh (2011) expuseram diversas espécies de cianobactérias e algas verdes a extrato de palha de cevada e obtiveram notável inibição de crescimento em algumas, incluindo as eminentemente perniciosas Microcystis aeruginosa e Anabaena flos-aquae. Martin e Ridge (1999) conduziram experimentos semelhantes e reportaram que, dentre as espécies testadas, quatro cepas de M. aeruginosa foram as mais suscetíveis.
O mecanismo químico por trás da atividade algistática da palha de cevada é desconhecido. A maior parte dos pesquisadores têm trabalhado com a hipótese de que a atividade inibitória provém da oxidação de extratos fenólicos liberados pela lignina encontrada em palha sob decomposição. Waybright et al. (2009) contestaram a sugestão de que o polifenol inibidor pode ser uma tanina, porquanto o extrato da palha reteve sua atividade após hidrólise; taninas são geralmente suscetíveis a hidrólise. Iredale et al. (2012) publicaram as primeiras evidências experimentais de que atividade microbiana pode ser a principal fonte dos componentes ativos de palha sob decomposição. O grupo de Iredale observou que amostras atípicas de fungos têm sido reportadas em experimentos com palha de cevada sob decomposição; ademais, Murray et al. (2010) mostraram que um pré-tratamento da palha com fungos lignolíticos pode reduzir o atraso entre a introdução da palha e a verificação de efeitos algistáticos.
Algumas algas parecem não ser afetadas por palha de cevada se esta não for aplicada de modo que a água seja consistentemente oxigenada. Isso foi verificado em escala laboratorial por Pillinger et al. (1994), que notaram que palha de cevada apodrecida somente produziu um efeito apreciável sobre o crescimento de Chlorella spp. se a aeração fosse aumentada nos últimos 7 dias do período de decomposição. De acordo com Boylan e Morris (2003), duas explicações para essa dependência de oxigênio podem ser que (1) oxigênio deve ser fornecido aos micróbios que decompõem a palha; ou (2) os compostos polifenólicos associados à atividade algistática da palha apresentam atividade superior quando oxidados. Todavia, Boylan e Morris (2003) mostraram que aplicação de extrato de palha em lagoas estagnantes em Iowa, EUA, não inibiu o crescimento de Cladophora spp.
2.4. Peróxido de hidrogênio
Peróxido de hidrogênio, H2O2, também já foi explorado como um produto algicida sustentável. H2O2 é um oxidante potente, capaz de desferir radicais hidroxila que danificam células e inibem atividade fotossintética por meio de perturbações ao fotossistema II; é altamente seletivo, afetando cianobactérias a concentrações 10 vezes menores do que aquelas que danificam algas verdes e diatomáceas (Drábková, 2007); possui ação rápida, de modo que um lago tratado com H2O2 pode ser seguro para natação e outras aplicações em um prazo não maior do que 3 dias (Matthijs et al., 2016); é decomposto a água e oxigênio ao invés de moléculas potencialmente tóxicas; finalmente, é acessível.
A maior sensibilidade de cianobactérias a H2O2 se comparadas ao fitoplâncton eucariota pode ser explicada pela sua resposta notavelmente distinta a estresse fotoquímico (Weenink, 2015; Matthijs, 2016). Algas eucariotas submetidas a luz intensa liberam H2O2 como um subproduto de fotossíntese. Ao nível molecular, exposição a luz intensa promove o aumento do fluxo de elétrons fotossintéticos, provocando a redução de oxigênio a ânion superóxido (reação de Mehler). Superóxido é um exemplo de espécie reativa de oxigênio, que é enzimaticamente convertido a H2O2 por superóxido dismutase. Finalmente, H2O2 é convertido a água e oxigênio por catalase ou, alternativamente, enzimas anti-ROS como ascorbato peroxidase reduzem H2O2 a água. Cianobactérias, por seu turno, não têm o aparato necessário para realizar uma reação de Mehler; estas reagem a estresse fotoquímico através de uma reação em que água é produzida diretamente mediante duas proteínas flavodiiron, sem H2O2 intermediário. Por consequência, níveis de estresse oxidativo são menores em células cianobacterianas do que em algas eucariotas, reduzindo a demanda por enzimas anti-ROS como ascorbato peroxidase, que inexiste na maioria das espécies de algas procariotas. Isso faz com que cianobactérias sejam mais sensíveis a H2O2 do que algas eucariotas e macrófitas.
Lusty e Gobler (2020) administraram H2O2 a amostras de água de 4 lagos eutróficos e, ao mensurar a diversidade microbiana após o tratamento, observaram que os gêneros de cianobactéria decresciam em suscetibilidade a H2O2 na ordem Planktothrix > Microcystis > Cylindrospermopsis. Lusty e seu colega notaram que Cylindrospermopsis spp. podem ser particularmente resistentes devido a seu arsenal de enzimas capazes de degradar H2O2, incluindo superóxido dismutase, catalase e ascorbato peroxidase. Microcystis spp., por sua vez, podem resistir a H2O2 porque formam colônias globulares de células envoltas em um muco dotado de polímeros extracelulares, os quais protegem as células intracoloniais contra agentes oxidantes como H2O2. Liu et al. (2017) verificaram que um tratamento de H2O2 a 5 mg/L foi insuficiente para perturbar grandes colônias de Microcystis, de modo que atividade fotossintética foi parcialmente restaurada 48 horas após a aplicação; uma dose de 20 mg/L, no entanto, mostrou-se capaz de suprimir o metabolismo das algas independentemente do tamanho das colônias. O emprego de altas doses de H2O2 pode assegurar a inibição das algas nocivas, mas aumenta o risco de lise celular e a subsequente liberação de toxinas intracelulares como microcistinas.
3. Dragagem de sedimentos
A oferta de fósforo interna pode sustentar o estado eutrófico de um lago por meses, ou mesmo anos, após a interrupção de fontes de poluição externas. A maior parte do suprimento interno de P provém dos sedimentos localizados no assoalho do lago; consequentemente, a dragagem de tais sedimentos deve favorecer projetos de restauração. Os efeitos de uma dragagem de sedimentos podem ser severos: Reddy et al. (2007) calcularam que a remoção dos 30 cm superiores do assoalho do lago Okeechobee, Flórida, EUA, equivaleram a mais de 120 anos de abastecimento “natural” de fósforo.
Um dos mais antigos casos de restauração por meio de dragagem de sedimentos concerne ao lago Trummen, Suécia (Björk, 2010). O lago havia sido usado para recreação e abastecimento até os anos 1920; deposição de águas residuais e outras formas de poluição levaram ao colapso hipertrófico do lago na década de 40. Embora a descarga de águas residuais tenha sido interrompida entre 1957 e 1958, o lago não se recuperou, mantendo transparência não melhor do que 20 cm no verão. Na década de 1960, gestores locais lançaram um projeto para restaurar o lago através de dragagem de sedimentos. A primeira metade dos 50 cm superiores do lago foi removida por dragagem de sucção em 1970, e a outra metade no ano seguinte; cerca de 400,000 metros cúbicos de sedimentos foram processados no total. Os benefícios foram imediatos: florações de algas desapareceram e uma comunidade diversa de fitoplâncton substituiu as monoculturas de Microcystis que predominavam anteriormente; o mexilhão de água doce Anodonta, que havia sido erradicado no período hipertrófico, voltou a colonizar o lago; o conteúdo de fósforo da água foi reduzido de 600
Sebetich e Ferriero (1997) descreveram o uso de dragagem de sedimentos no lago Kampfe, Nova Jersey, EUA, e compararam suas características pós-intervenção com o lago Glen Wild, um lago vizinho que não foi dragado mas tinha história de desenvolvimento semelhante e, portanto, poderia ser usado como ‘controle.’ O lago Kampfe exibiu condições superiores, incluindo uma concentração de fósforo média ao longo da coluna d’água igual a 20.9
Figura 2. Perfis de concentração de fósforo nos lagos Kampfe e Glen Wild. As datas referem-se ao ano de 1993.

Experimentos de laboratório indicaram que, como no caso de outras técnicas de restauração, o sucesso de uma dragagem de sedimentos depende indiretamente do controle de fontes externas de fósforo (Kleeberg e Kohl, 1999; Zhong et al., 2007). Isso é especialmente verdadeiro no longo prazo; Liu et al. (2016) avaliaram os efeitos de dragagem sobre o carregamento interno de N e P ao longo de 15 anos no lago Wuli, um lago urbano na cidade de Wuxi, China, e verificaram que, 18 meses após a intervenção, a taxa de liberação de fósforo solúvel reativo (SRP) havia retornado a níveis pré-dragagem. Liu e seu grupo acreditam que isso foi uma consequência da contínua poluição do lago por fontes externas, mesmo após a dragagem.
Li et al. (2020) estudaram a dinâmica pós-dragagem de um lago aquicultural na China e concluíram que essa técnica de restauração não é um remédio definitivo contra eutroficação. Especificamente, Li e seu grupo concluíram que, após a dragagem, a redeposição de sedimentos naturais promove o enterro de fósforo biodisponível e a transformação de P inerte a P ativo, aumentando o risco de liberação interna de P e, no longo prazo, promovendo o retorno a condições eutróficas. Li e seu grupo afirmam que, a fim de minimizar a liberação de P ativo, projetos de dragagem devem incorporar técnicas de gerenciamento in situ como o plantio de macrófitas aquáticas, com o intento de aumentar a espessura da camada de óxido dos sedimentos superficiais e evitar a resuspensão de sedimentos.
4. Aeração/oxigenação hipolimnética
Aeração ou oxigenação hipolimnética (AH/OH) são técnicas utilizadas para manter ou aumentar a concentração de oxigênio na camada inferior de um lago por meio da injeção de ar ou oxigênio. Conforme mencionado na seção 3, a reserva de fósforo nos sedimentos de um lago pode ser muitas vezes maior do que a concentração suspensa na água, de modo que a oferta de P é altamente dependente da dinâmica sedimento-água (Søndergaard et al., 2003). Sabe-se que sedimentos liberam mais fósforo quando estão sob condições anóxicas. Portanto, níveis superiores de oxigênio no hipolímnio podem reduzir a liberação de fósforo na interface sedimento-água e aumentar a retenção de fósforo, contribuindo assim para a redução do nível trófico do corpo d’água.
O uso de aeração e oxigenação hipolimnética tem produzido resultados bons e ruins. Schauser e Chorus (2007) descreveram a restauração do lago Tegel, Alemanha, através da instalação de 15 aeradores juntamente com uma redução sistemática na deposição externa de fósforo. Utilizando um modelo matemático, Schauser e sua colega concluíram que o uso dos aeradores não teria tido impacto sobre a restauração se não fosse complementado por uma redução de P externo. Liboriussen et al. (2009) reportaram que, em 5 lagos dinamarqueses que foram oxigenados artificialmente por 4 a 20 anos, houve apenas aprimoramentos mínimos em termos de conteúdo de oxigênio no hipolímnio, com concentrações de O2 no verão não maiores do que 2.2 mg L–1; ademais, as melhoras restringiram-se aos primeiros meses de estratificação. Moore et al. (2012) documentaram o uso de oxigenação hipolimnética no lago Newman, Washington, EUA, que, com profundidade média de 5.6 m, é um dos lagos mais rasos em que a técnica foi utilizada até o momento. O uso de OH foi capaz de prevenir anoxia na maior parte dos anos de observação, mas os oxigenadores tiveram de operar sob capacidade total (operação sob capacidade parcial não mostrou-se suficiente) e a interrupção do suprimento de O2 em qualquer estação do ano foi rapidamente seguida pelo esgotamento do oxigênio hipolimnético.
É importante observar que aeração e oxigenação hipolimnética, embora similares, utilizam gases diferentes – ar ou oxigênio puro – e, portanto, podem produzir efeitos distintos. Austin et al. (2019) descreveram o uso de AH e OH em dois lagos no estado de Minnesota, EUA, no decurso de 30 anos; o uso de oxigenação produziu resultados superiores a aeração não apenas em termos de redução de anoxia, mas também em termos de oxidação de ferro e manganês, supressão de fósforo interno, redução de fósforo total superficial e redução de extremos de concentração de clorofila-a.
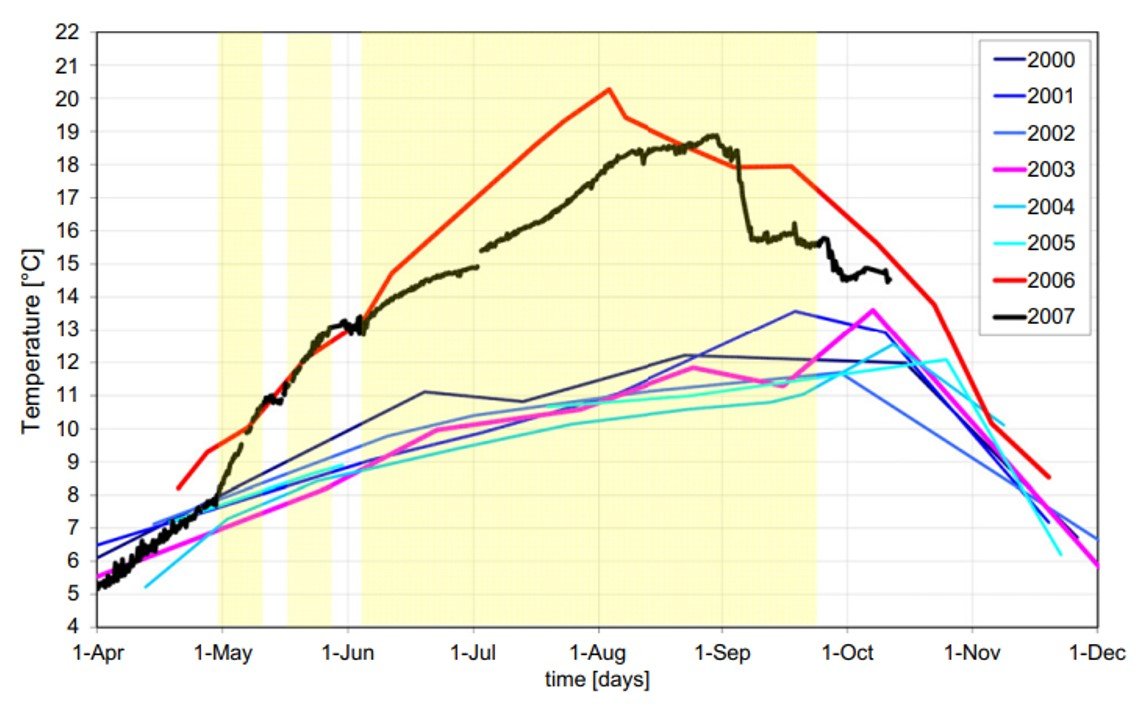
O uso de AH e OH é comumente atrelado a aumentos na temperatura do hipolímnio, o que pode enfraquecer a estratificação e conduzir a um overturn prematuro no fim do verão. Trabalhando com dados do lago Serraia, Itália, Toffolon et al. (2013) reportaram que temperaturas mensuradas nas águas mais profundas aumentaram até 9oC após a introdução de um aparato de oxigenação (Figura 3). Toffolon e seu grupo utilizaram uma análise semiquantitativa para mostrar que reações exotérmicas associadas à degradação de matéria orgânica não poderiam justificar completamente tal variação. Para eles, o aumento na temperatura do hipolímnio deve ser atribuído à circulação de massas d’água e à difusão vertical provocada por injeções de água no fundo do lago; ou seja, o aumento de temperatura tem relação direta com o equipamento de oxigenação.
Figura 3. Temperaturas observadas no hipolímnio do lago Serraia, Itália, entre 2000 e 2007. A oxigenação artificial foi iniciada em 2006; observe as temperaturas particularmente elevadas para os anos de 2006 e 2007.
5. Biomanipulação
Altas densidades de peixes planctívoros-bentívoros podem impedir o crescimento de macrófitas aquáticas e reduzir populações de zooplâncton, favorecendo a dominância do fitoplâncton sobre o ecossistema lacustre. Uma hipótese lógica, portanto, é que a remoção parcial ou completa de tais peixes pode favorecer o zooplâncton, aumentar a pressão de herbivoria e promover a redução de algas nocivas. Isso é um exemplo de biomanipulação, isto é, uma modificação deliberada em uma comunidade ecológica e suas interações. Esquemas de biomanipulação de vários tipos têm sido utilizados como uma técnica de restauração de lagos nos últimos 30 anos. Devido à dificuldade em controlar populações de peixes em grandes corpos d’água, a maior parte das iniciativas do gênero tem sido executada em lagos rasos e lagoas.
Como mencionado acima, a abordagem de biomanipulação mais comum consiste na remoção de peixes zooplanctívoros e bentívoros. Uma redução na biomassa de tais peixes é geralmente acompanhada por uma diversidade de efeitos ecológicos incluindo redução na biomassa de fitoplâncton, favorecimento de zooplâncton grandes, maior transparência e maior proporção de peixes piscívoros. É importante observar que a biomanipulação somente produz os resultados desejados se a remoção de peixes for suficientemente expressiva. Uma remoção de 200 kg ha–1 ao longo de um período de 3 anos foi recomendada por Olin et al. (2006), que basearam essa estimativa em suas experiências com lagos finlandeses; Jeppesen e Sammalkorpi (2002) propuseram que, em lagos temperados rasos, a massa C de peixes a ser removida para obter aprimoramentos na qualidade da água é dada pela lei de potência C = 6.9 
Uma segunda modalidade de biomanipulação em ambientes lacustres é a introdução de espécies piscívoras. A lógica, nesse caso, é que os peixes piscívoros consumirão as espécies zooplanctívoras e bentívoras, produzindo uma cascata trófica semelhante à obtida se tais espécies fossem simplesmente removidas. Piscívoros comumente utilizados incluem picão-verde e achigã nos EUA e lúcio na zona temperada europeia. Infelizmente, a experiência com biomanipulação mediada por piscívoros tem produzido resultados decepcionantes. Drenner e Hambright (2002), revisando 17 estudos, reportaram que a maioria não indicou a existência de uma relação entre adição de piscívoros e biomassa de fitoplâncton. Alguns gestores têm combinado introdução de piscívoros com remoção de plancto-bentívoros, mas mesmo em tais casos questionou-se o valor adicional da primeira técnica em relação ao uso da última (Triest et al., 2016).
A principal limitação de operações de biomanipulação, provavelmente, é seu efeito limitado a longo prazo, porquanto vários estudos de caso relatam um retorno de águas túrbidas e populações substanciais de zooplanctívoros dentro de 5 a 10 anos. O mais detalhado estudo acerca de experiência com biomanipulação no longo prazo é Søndergaard et al. (2008), que estudaram 36 lagos eutróficos dinamarqueses nos 10 – 15 anos subsequentes à remoção de peixes. Nos primeiros 6 – 8 anos, Søndergaard e seu grupo notaram aprimoramentos significativos nas variáveis de interesse, incluindo maior profundidade de Secchi e menor concentração de clorofila-a, na maior parte dos lagos. Após aproximadamente 10 anos, no entanto, a maior parte das variáveis tenderam a retornar a valores pré-intervenção. Reduções na abundância de cianobactérias mostraram-se estatisticamente significativas apenas nos 3 – 7 anos iniciais.
Uma vez que intervenções de biomanipulação são frequentemente executadas uma única vez, há necessidade de mais investigações acerca de remoções de peixes reiteradas. Uma contribuição recente, novamente oriunda da Dinamarca, é Søndergaard et al. (2017), que estudaram 30 anos de dados relativos à restauração do lago Væng. O lago foi submetido a remoção de peixes duas vezes, primeiramente em 1986 – 1988 e novamente em 2007 – 2009. Søndergaard e seu grupo mostraram que a segunda biomanipulação replicou os efeitos químicos e biológicos da primeira, ainda que 30% menos peixes tenham sido removidos na segunda intervenção. Portanto, parece que remoções subsequentes de peixes podem ser mais fáceis de implementar e igualmente eficazes.
6. Ultrassom
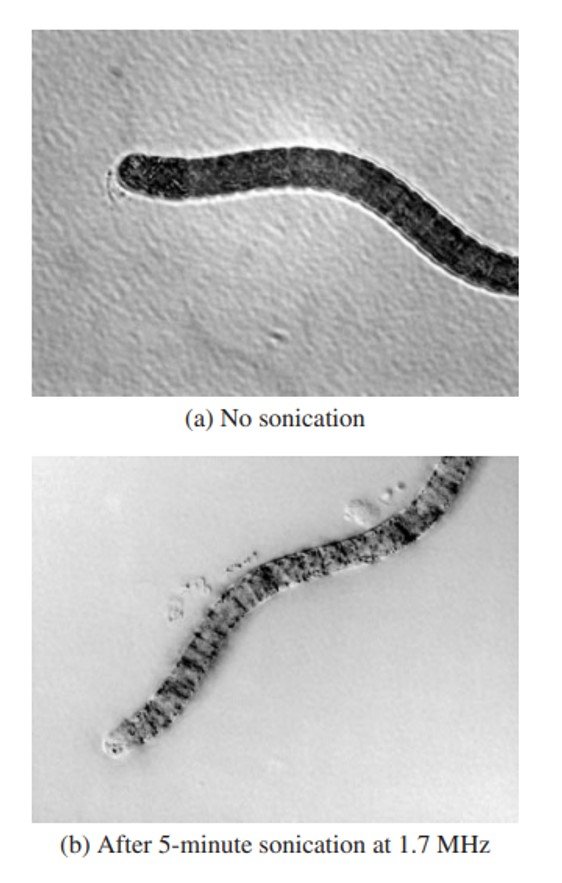
O uso de radiação ultrassônica mostrou-se eficaz no controle de Spirulina e espécies cianobacterianas dotadas de vacúolos. Radiação ultrassônica em ambientes aquáticos produz uma série de ciclos de compressão e rarefação que geram milhões de bolhas de cavitação; a implosão dessas bolhas produz temperaturas da ordem de 5,000oC e pressões da ordem de 500 atm, além de formar radicais livres. A sonicação de cianobactérias inibe seu crescimento devido à ruptura de seus vacúolos diante de efeitos cavitacionais, interrupção de atividade fotossintética e perturbação do ciclo celular (Figura 4). Zhang et al. (2006) notaram que a remoção de algas por sonicação em escala laboratorial segue uma reação de primeira ordem.
O controle de algas por sonicação é dependente de parâmetros como frequência, intensidade e período de exposição. Hao et al. (2004) testaram fontes ultrassônicas a 20 kHz, 200 kHz e 1.7 MHz e reportaram que o efeito inibitório era mais eficaz a 200 kHz. Joyce et al. (2010) usaram fontes a 20, 40, 580, 864 kHz e 1.146 MHz sob várias intensidades por 30 min e verificaram que, embora a fonte de 864 kHz tenha produzido a maior redução de algas, a fonte de 580 kHz era mais eficiente (com eficiência definida como porcentagem da população algal inativada dividida pela intensidade da fonte). Sonicação sob frequências elevadas requer intensidades sonoras mais elevadas, e consequentemente mais potência elétrica, para obter cavitação. Sonicação sob potências maiores tem sido associada com remoção mais ampla de algas, mas é acompanhada de lise celular e, consequentemente, acumulação de toxinas extracelulares como microcistinas (Zhang et al., 2006; Park et al., 2019). Da mesma forma, um maior período de exposição é acompanhado de maior exposição a efeitos cavitacionais e lise celular. Todavia, sabe-se que ultrassom é eficaz na remoção de toxinas aquáticas, e alguns pesquisadores têm reportado que a sonicação prolongada de M. aeruginosa, ao menos em condições de laboratório, pode reduzir o nível de substâncias nocivas ao invés de aumentá-lo (Rajasekhar et al., 2012). A questão de que, no campo, a aplicação prolongada de ultrassom conduz a um acúmulo ou redução de toxinas algais continua sendo debatida, mas as evidências presentes favorecem a primeira hipótese.
Figura 4. Uma célula de alga filamentosa (a) antes de sonicação e (b) após 5 minutos de sonicação a 1.7 MHz.
Referências
A lista de referências pode ser encontrada no meu post original.